








Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkActa Biológica Colombiana
versión impresa ISSN 0120-548X
Acta biol.Colomb. v.14 supl.1 Bogotá dic. 2009
DARWIN Y LA PARADOJA DE LAS ISLAS VACÍAS
Darwin and the Empty Island Paradox
JUAN FELIPE BLANCO1, Ph. D. 1Grupo de Limnología Básica y Experimental, Instituto de Biología. Universidad de Antioquia. Medellín, Colombia. blanco@exactas.udea.edu.co
Presentado 21 de septiembre de 2009, aceptado 26 de noviembre de 2009, correcciones 19 de mayo de 2010.
RESUMEN
Aunque se conoce el interés y fascinación de Darwin por la naturaleza marina a través de sus tratados sobre arrecifes coralinos, islas oceánicas y balanos, poco se comenta sobre los múltiples ejemplos dulceacuícolas que él utilizó para plantear la -teoría de la migración- dentro de -El origen-. Ésta plantea que la fauna y la flora se -dispersan- desde los continentes hacia las islas oceánicas. Sin embargo, en la islas muy alejadas de los centros de origen la probabilidad de colonización es muy baja y por lo tanto solo se encuentra una fracción de la biota continental, pudiendo considerarse en algunos casos -vacías-. Aunque las corrientes de agua de dichas islas también se consideran -vacías-, Darwin propuso un mecanismo para explicar la presencia de peces -dulceacuícolas- en las mismas, y por lo tanto no podrían considerarse -vacías-, lo cual plantea una paradoja. El mecanismo planteado por él coincide con el ciclo de vida conocido actualmente como diádromo en el cual los individuos de muchas especies de peces, camarones y gasterópodos requieren de aguas marinas y dulces para completar su desarrollo. La diadromía es una convergencia evolutiva que apareció en diferentes linajes a partir de ancestros tanto marinos como dulceacuícolas. En este ensayo se discute la evolución de la fauna dulceacuícola insular, sus implicaciones ecológicas y evolutivas, y algunos modelos experimentales. Finalmente, se discute sobre los impactos de las represas en islas tropicales y subtropicales sobre las especies diádromas.
Palabras clave: biogeografía insular, diadromía, quebradas insulares, teoría de la migración.
ABSTRACT
Although Darwin’s fascination and interest on marine nature are well known due to his treatises about coral reefs, oceanic islands, and barnacles, little is commented about the many -freshwater- examples that he provided for supporting -The Migration Theory- in his book -The Origin of the Species-. According to this theory, faunas and floras disperse from continents to oceanic islands. However, those islands located at a great distance from the center of origin are very unlikely to be colonized by immigrants, and therefore only a fraction of the continental biota is represented in these islands, and they could be considered as empty. Streams in these islands could also be empty, however, Darwin stated a mechanism to explain the presence of freshwater fish in these streams, unaware of an emerging paradox. This mechanism is consistent with the current definition of a diadromous life cycle observed in fish, shrimp and snails that rely on migrations between marine and freshwaters to complete their development. This life cycle is an evolutionary convergence among unrelated lineages that evolved from both marine and freshwater ancestors. This essay discusses the evolution of insular freshwater faunas, their implications as ecological and evolutionary forces, and some experimental models. A final discussion is presented on the impacts of dams on diadromous fauna in tropical and subtropical islands.
Key words: Diadromy, island biogeography, island streams, migration theory.
PARADOJA
- Aserción inverosímil o absurda, que se presenta con apariencias de verdadera.
- Figura de pensamiento que consiste en emplear expresiones o frases que envuelven contradicción (Diccionario de la Real Academia de la Lengua Española).
Qué gran hecho –qué misterio de misterios– es la aparición de nuevos seres sobre la tierra Charles Darwin (diario del Beagle).
There is a striking parallelism in the laws of life throughout time and space: laws governing the succession of forms in past times being nearly the same with those governing at the present time the differences in different areas Edward Forbes (naturalista británico: 1815-1854), citado por Darwin en El origen.
INTRODUCCIÓN
Con la conmemoración de los 150 años de la publicación de El origen de las especies en 1859 (Darwin, 1859; o simplemente El origen como el mismo Charles Robert Darwin y sus amigos se referían a esta obra), se han revisado nuevamente sus aportes a aquello que posteriormente se denominó la teoría evolutiva. Adicionalmente, la conmemoración del bicentenario del natalicio de Darwin ha servido para escudriñar su vida personal y sobre todo su obra completa, la cual se encuentra disponible en línea (Darwin, 1839). Sin embargo, hay aspectos de su vida y aportes científicos que todavía no han sido explorados en profundidad. El objetivo de este artículo es precisamente mostrar a través de El origen y otras de sus obras, que Darwin sintió una profunda atracción por los sistemas dulceacuícolas (los cuales aparentemente conocía bien) y también exponer cómo, sin proponérselo, planteó las bases de lo que he denominado La paradoja de las islas vacías.
La obra de Darwin incluye varios tratados relacionados con la naturaleza marina, pero también en algunas secciones de sus obras diserta sobre los -seres orgánicos- dulceacuícolas y su -entorno- (para no utilizar términos como hábitat y ecosistema que fueron propuestos en los albores del siglo veinte). El Darwin -marino- se muestra claramente en La zoología del viaje del H. M. S. Beagle (1838-1843), La estructura y distribución de los arrecifes de coral (1842), Observaciones geológicas acerca de las islas volcánicas visitadas durante el viaje del H. M. S. Beagle (1844) y Una monografía sobre la subclase Cirripedia (cuatro volúmenes editados entre 1851 y 1854 en los que abarca los balanos vivientes y fósiles de dos familias; todas disponibles en línea: Darwi, 1838-1843). En La zoología del Beagle quiero detenerme en un volumen sobre -Peces- (Parte 4, 1842) que fue editado por el reverendo Leonard Jenyns. Aunque Darwin coeditó este y otros de los volúmenes de la Zoología del Beagle, y en algunos de ellos contribuyó escribiendo secciones enteras, en el volumen sobre los peces solo se transcribieron muchas de las notas que se encontraban en las etiquetas de colección de los especímenes. En este trabajo se incluyeron descripciones de muchos especímenes de agua dulce de las costas de Suramérica y Nueva Zelanda. Por otra parte, Darwin dedicó dos capítulos de El origen a la -distribución geográfica- en los cuales utilizó muchos ejemplos de biota marina y algunos de biota dulceacuícola.
Las obras anteriormente citadas se basan completamente en las experiencias vividas y muestras recolectadas por Darwin durante el viaje alrededor del mundo a bordo del Beagle entre 1832 y 1836. Aunque mucho se ha hablado del paso por la costa suramericana y, particular-mente, por las Islas Galápagos, El Beagle también visitó las islas Falklands, varias islas del océano Pacífico central, Nueva Zelanda, el océano Índico y el océano Atlántico central (Diario de abordo Darwin, 1839). Por esta razón, las teorías expuestas por Darwin incluyen numerosos ejemplos insulares. Por lo tanto, el primer objetivo de este ensayo es resaltar la importancia que tuvieron las islas para el desarrollo de la teoría evolutiva y sus implicaciones para la limnología insular. Para ello he traducido apartes de El origen tratando de mantener el sentido más fiel de la versión original en inglés. El segundo objetivo es describir la diadromía como evento evolutivo convergente en la fauna lótica insular y hacer una revisión de las implicaciones de esta estrategia de vida para la ecología de las comunidades y de los ecosistemas insulares.
SOBRE LA DISPERSIÓN Y LA BIOTA INSULAR
En El origen, Darwin incluye dos capítulos (11 y 12) sobre la -distribución geográfica- para proponer lo que en el texto denominó La teoría de la migración. En la exposición de argumentos a favor de la misma, que utilizó exponiendo muchos ejemplos de biota marina y algunos de biota dulceacuícola. En el capítulo 11 plantea:
La similitud o disimilitud en los habitantes de varias regiones no puede ser atribuida a sus condiciones climáticas y otras actualmente predominantes- -...cada especie ha sido ‘producida’ dentro de un área, y ha migrado tanto como ha podido. Una isla volcánica...ha probablemente recibido de él (un continente), en el curso del tiempo, unos pocos colonos, y sus descendientes, aunque modificados, estarían aun plenamente relacionados por herencia con los habitantes del continente. La gran importancia de las barreras aparece cuando miramos la migración; como lo es tiempo para el lento proceso de modificación por selección natural. El grado de disimilitud dependerá de la migración de las formas de vida más dominantes desde una región hacia otra, siendo esto más o menos fácil en períodos más o menos remotos;...
En los anteriores planteamientos Darwin propone que la separación espacial de las regiones del mundo es un mecanismo tan importante como el tiempo, que puede explicar la presencia de especies en un lugar determinado, lo mismo que su similitud con la de otras regiones. Así como la descendencia con modificación podía explicar la -aparición- de especies en una región, la migración (-...cuando el clima fue diferente pudo haber una ‘vía’ para la migración, pero ahora es impasable-) también podía explicar la -aparición- de las mismas especies en regiones actualmente separadas por barreras. Esto suponía dos condiciones. La primera, es que las especies habían migrado en el pasado desde un centro de origen hacia otra región. La segunda, es que actualmente no era posible para los individuos de las mismas especies migrar desde el centro de origen debido a que alguna barrera lo impedía. Aquí también es clara la alusión a las islas volcánicas como ejemplo. Más adelante Darwin explica los mecanismos particulares a través de los cuales los organismos migraron entre regiones. En la sección -medios de dispersión- explica que en los tiempos benignos las especies pudieron migrar desde sus centros de origen continentales hacia las islas oceánicas a través de puentes de tierra (land bridges) formados cuando el nivel del mar estuvo más bajo. En ausencia de estos puentes, las especies también pudieron haber tomado ventaja de la ubicación consecutiva de varias islas, y por lo tanto pudieron haber -saltado- desde su centro de origen hasta las islas oceánicas (stepping stones). Sin estos puentes y -caminitos de piedras- la dispersión era bastante improbable según Darwin, debido a que para el caso de la biota terrestre el mar representa una barrera impasable. Por ejemplo, mientras los animales mamíferos y reptiles deberían poseer una gran capacidad de natación, las plantas y los animales invertebrados debían soportar los efectos negativos de la salinidad como adultos, o tener semillas o huevos, respectivamente, con capacidad de permanecer en estado de latencia durante largos períodos y aun así poder germinar o eclosionar. Por esta razón, Darwin pensaba que los dos mecanismos expuestos inicialmente (i.e. los puentes y los caminos) solo podían explicar la dispersión hacia islas cercanas a los continentes u otras más grandes, pero no hacia las islas oceánicas ubicadas en el centro de los océanos. De hecho, él notó que algunos grupos de animales (e.g. mamíferos no voladores y anfibios) y plantas (e.g. de semillas grandes) estaban ausentes en las islas oceánicas. Esto lo sintetiza en el siguiente enunciado:
La naturaleza y las proporciones relativas de los habitantes de las islas oceánicas me parecen de esta manera opuestas a la creencia de que estuvieron conectadas a los continentes.
Dada la improbabilidad de formar puentes o -caminitos- entre los continentes y las islas oceánicas, Darwin pensó que la dispersión hacia éstas era de carácter accidental u ocasional. En el capítulo 11 cita varios ejemplos de cómo plantas y animales pueden viajar durante muchos días
o meses sobre islotes de tierra o troncos que son arrastrados desde las desembocaduras de los ríos o las costas (proceso conocido actualmente como rafting). También explicó cómo semillas de plantas y huevos de caracoles terrestres podían viajar con el fango en las patas de las aves migratorias (fenómeno conocido como foresia). Finalmente, plantea que los huracanes y tormentas pueden transportar con sus fuertes vientos a organismos pequeños, a lo cual denominó dispersión catastrófica. De todas formas, aunque los organismos pudiesen llegar a islas oceánicas suficientemente alejadas de los continentes por medio de dispersión ocasional, accidental o catastrófica, éstos debían, de todas formas, ser resistentes a las inclemencias ambientales y tener la capacidad de germinación tardía, según lo resalta el propio Darwin. Obviamente muchos patrones permanecieron sin explicación para él como lo plantea a continuación:
Indudablemente existen muchos casos, en los cuales no podemos explicar cómo la misma especie ha pasado de un punto a otro. Esta visión..., no difiere mucho... de la elaborada recientemente por el Sr. Wallace en un ingenioso artículo.
Las ideas sobre la importancia que tiene la migración de plantas y animales para la colonización de las islas oceánicas fueron el fundamento y la inspiración para lo que a mediados del siglo veinte se formalizara como la teoría de la biogeografía de Islas (MacArthur y Wilson, 1967). Esta teoría plantea que la tasa de inmigración en las islas es función inversa del aislamiento.
LA PARADOJA DE LAS ISLAS VACÍAS Aunque Darwin no planteó explícitamente una paradoja con respecto a la biota insular, existen dos líneas de argumentación dentro del capítulo 12 que son aparentemente contradictorias y que se explicarán más adelante. A esta aparente contradicción es lo que he denominado la -paradoja de la islas vacías-. Adicionalmente, debo precisar que esta paradoja se limitada exclusivamente a los ríos y quebradas de las islas. Por definición, una paradoja implica a) una aserción inverosímil con apariencia de verdad y b) una contradicción. Se excluyen los sistemas lénticos insulares porque la evidencia ha mostrado que efectivamente tienen menos especies e individuos que sus equivalentes continentales, y por lo tanto la teoría es consistente con la evidencia de campo.
Darwin anota en el capítulo 12 que las -producciones dulceacuícolas- (biota) de los lagos y ríos posee una naturaleza insular debido a que están aisladas por masas de tierra. Adicionalmente, plantea que dicha biota será aun más distinta entre regiones separadas por los océanos (-una barrera aun más impasable-). Pero menciona que, contrariamente, durante su viaje lo impresionó el gran parecido que tenía la biota acuática del Brasil con la de Inglaterra:
Bien recuerdo, cuando estuve recolectado en las aguas dulces de Brasil, sentí mucha sorpresa debido a la similitud de los insectos, conchas, etc., y la disimilitud de los ‘seres’ terrestres que los rodeaban, al compararlos con aquellos de Britania.
Ante este hecho propone la siguiente hipótesis:
Pero esta capacidad para moverse ampliamente que tienen los animales de agua dulce... resultó muy útil para ellos, para las migraciones cortas y frecuentes entre charcas o entre ríos; y la amplia dispersión pudo haber seguido a esta capacidad como consecuencia.
De acuerdo a esto, entonces las aguas dulces de las islas oceánicas deberían poseer una fauna similar a la de los continentes. Darwin no ofrece una respuesta basada en los insectos, pero se podría utilizar el argumento por él planteado en el capítulo 11 sobre los mecanismos de dispersión de la biota terrestre para obtener una respuesta. Los insectos acuáticos con estados adultos terrestres o con poca capacidad de vuelo pudieron colonizar las islas oceánicas en tiempos en que existieron puentes o -caminitos- con los continentes. La distancia, sin embargo, seguiría siendo una barrera para ellos, y por lo tanto la colonización de las islas más alejadas estaría limitada a aquellas especies con estados adultos de gran capacidad de vuelo o a la dispersión accidental, ocasional o catastrófica. Por lo tanto, los ríos y quebradas de las islas oceánicas podrían estar relativamente -vacíos- en términos de entomofauna.
Pero Darwin sí aborda el problema de la dispersión de los peces de agua dulce. Esto parte del interés y conocimiento aparentes que de ellos tenía. Por ejemplo, en el volumen dedicado a los peces dentro de -La Zoología del Beagle- se describen especímenes de varias familias primarias de agua dulce (Poecillidae, Lebiasinidae, Cyprinidae y Siluridae). También se relacionan familias dulceacuícolas (Salmonidae) o marinas (Gobiidae) cuyos miembros tienen una gran capacidad de migración. Considero que Darwin aprovecha este conocimiento para su argumento sobre la distribución y los mecanismos de dispersión de los peces. Con respecto a los patrones anota que -la misma especie de pez no puede estar en las aguas de continentes distantes-. Por analogía, la misma especie tampoco estaría en el continente y en las islas oceánicas, por lo tanto éstas también estarían vacías en términos de peces dulceacuícolas.
Pero resalta que en el mismo continente hay distribuciones amplias y -caprichosas-. Las similitudes y diferencias de la ictiofauna de dos ríos, según él, podrían ser explicadas de varias maneras. Por una parte, habla del transporte accidental, citando el ejemplo de los torbellinos de viento en la India que transportan peces vivos y huevos. Pero se inclina más por -la dispersión reciente a través de los cambios en el nivel de la superficie de la tierra causando confluencias entre los ríos-. También habla de las crecientes. En la escala de tiempo geológico explica:
Con respecto a las especies de peces de agua dulce que existen en puntos muy distantes del mundo, sin duda hay muchos casos en los cuales no hay explicación; pero algunos peces de agua dulce pertenecen a formas muy ancestrales, y en tales casos habrían tenido un tiempo suficientemente amplio para (experimentar) grandes cambios geográficos, y consecuentemente el tiempo y los medios para tal migración.
Esta descripción corresponde a lo que actualmente se conoce como vicarianza por deriva continental o por separación de masas de tierra por procesos tectónicos (Brown y Lomolino, 1998). Debido a la separación entre masas de tierra, los linajes dulceacuícolas evolucionados después del evento vicariante estarían restringidos a una u otra masa, a menos que se diese un proceso de dispersión reciente.
Darwin plantea una elaborada hipótesis para explicar la -dispersión reciente- de peces de agua dulce a través del mar:
En segundo lugar, los peces de aguas saladas, pueden con cuidado ser acostumbrados lentamente a vivir en aguas dulces; y de acuerdo a Valenciennes, no hay un solo grupo de peces confinado exclusivamente a aguas dulces, así es que nos imaginamos que un miembro marino de un grupo de agua dulce pudo viajar a través de las costas del mar, y subsecuentemente modificarse y adaptarse a las aguas dulces en tierras distantes.
Hasta el momento no he leído alusión a esta hipótesis de Darwin en la literatura limnológica insular, por lo cual creo que ha pasado inadvertida. Sin embargo, ésta describe una estrategia de vida conocida como la diadromía la cual ha sido ampliamente estudiada desde la segunda mitad del siglo veinte (McDowall, 1988) y que será abordada más adelante. De acuerdo a esta última hipótesis planteada por Darwin las islas oceánicas deberían estar dominadas por especies de peces de linajes en los que alternativamente se dio el salto al agua marina y posteriormente a agua dulce.
Darwin concluye -Sobre los habitantes de las islas oceánicas-:
Las especies de todas las clases... son pocas en número comparadas con aquellas en áreas continentales iguales:... ... son algunas veces deficientes en ciertas clases, y su lugar es aparentemente ocupado por otros habitantes; ... ...en las islas Galápagos..., 2 de 11 especies de aves marinas, (pocas especies) son peculiares:...
Muchas pequeñas particularidades notables podrían anotarse...
Aunque él no hace una síntesis de los ejemplos planteados para las aguas dulces insulares, utilizando la misma lógica, se puede concluir que las especies de insectos y otros invertebrados acuáticos y de peces de agua dulce: 1. -... son pocas en número:...-, 2. -... son algunas veces deficientes en ciertas clases...-, 3. -Muchas pequeñas particularidades notables podrían anotarse...- y 4. -... (pocas especies) son peculiares:...- Las tres primeras conclusiones apoyan la hipótesis de las islas vacías. Entonces, ¿cuál es la paradoja? De acuerdo a la última hipótesis los ríos, quebradas, riachuelos y arroyos insulares no están vacíos porque están colonizados por peces que han evolucionado a partir de linajes marinos o dulceacuícolas con transiciones marinas. De hecho las evidencias actuales apoyan este último planteamiento (Fig. 1). Por lo tanto, la paradoja consiste en que aparentemente las aguas corrientes insulares están vacías, pero esto se contradice por la abundancia de algunos peces, camarones y caracoles.

¡SÍ, LAS ISLAS ESTÁN VACÍAS! ¡NO, ESTÁN LLENAS!
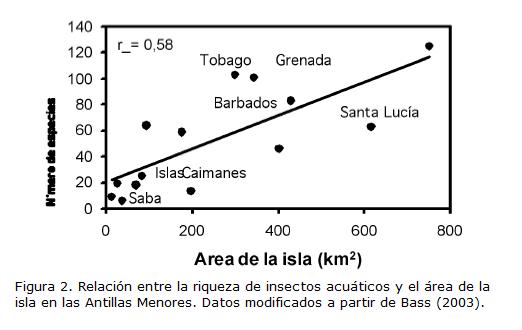
Los estudios disponibles permiten establecer que el aislamiento, la edad de la isla y el tamaño son variables determinantes de la riqueza de insectos lóticos, es decir de las aguas corrientes. Por ejemplo, en las quebradas del archipiélago de Hawaii (ubicadas a >4.000 km del continente más cercano) no existen Ephemeroptera, Plecoptera y Trichoptera nativos dadas sus limitaciones de vuelo, y se cree que los Odonata han radiado a partir de un ancestro común (Brasher, 2003). De manera equivalente, en el archipiélago de la Polinesia (ubicadas a >4.000 y >6.000 km de Australia y Suramérica, respectivamente) los simúlidos (Diptera) han sufrido una amplia radiación a partir de un ancestro común (Craig, 2003). El número de especies de simúlidos varía con la edad geológica de estas islas oceánicas volcánicas. En las islas muy jóvenes (menos de trescientos mil años) no ha transcurrido el tiempo suficiente para que se hayan establecido especies, mientras que en las islas muy viejas (más de tres millones de años) ya se han erosionado los hábitats preferidos (i.e. cascadas) por estos insectos. La mayor riqueza se encuentra en las islas con edades intermedias en las cuales ha ocurrido colonización por parte de un ancestro común y una posterior especiación, en los diver-sos hábitats disponibles. En las Antillas Mayores y Menores están ausentes los órdenes Plecoptera y Megaloptera, pero además la similitud de especies es baja entre islas (Bass, 2003). Además, en esta región se ha encontrado que la riqueza está relacionada directamente con el área (Fig. 2) y elevación máxima de la isla, patrones que coinciden con los hallazgos de Vinson y Hawkins, 2003, quienes utilizaron una base de datos global. Los insectos terrestres de las islas han sido objeto de más estudio que los acuáticos (Gillespie y Roderick, 2002).
En los sistemas lóticos insulares, la pobreza y poca abundancia de insectos, otros invertebrados y peces primariamente dulceacuícolas (sensu Myers, 1949; McDowall; 2004) contrasta con la gran abundancia y biomasa de peces, camarones y gasterópodos con ciclos de vida diádromos, es decir que tienen unos estados de desarrollo en el mar y otros en las aguas dulces (Smith et al., 2003; Covich, 2006). Por ejemplo, en la isla de Puerto Rico, Blanco y Scatena, 2005, han documentado hasta 7.000 individuos/m2 de una especie de caracol diádromo (Neritina virginea). En algunas islas del pacífico central la fauna lótica llega a estar compuesta completamente por fauna diádroma (March et al., 2003; Covich, 2006; Benstead et al., 2009).
LA DIADROMIA COMO ESTRATEGIA EVOLUTIVA CONVERGENTE EN LAS QUEBRADAS INSULARES
La descripción hecha por Darwin acerca de la dispersión de peces dulceacuícolas a través del mar coincide con la estrategia diádroma. Es posible que Darwin tuviese conocimiento de primera mano sobre dicha estrategia dado que capturó varios peces en Nueva Zelanda (La zoología del Beagle), donde la mayoría de la ictiofauna dulceacuícola es diádroma (McDowall, 1988; McDowall, 1993a). Entre ellos capturó varios salmónidos, los cuales comparten dicho comportamiento con los salmónidos norteamericanos y británicos. También capturó a Eleotris gobioides el cual es diádromo. La descripción de éste en Los peces del Beagle dice lo siguiente:
This species was taken by Mr. Darwin in fresh-water, in the Bay of Islands, New Zealand. It so well accords with the description of the E. gobioides in the ‘Histoire des Poissons’, that I conceive there can be no doubt of their identity.
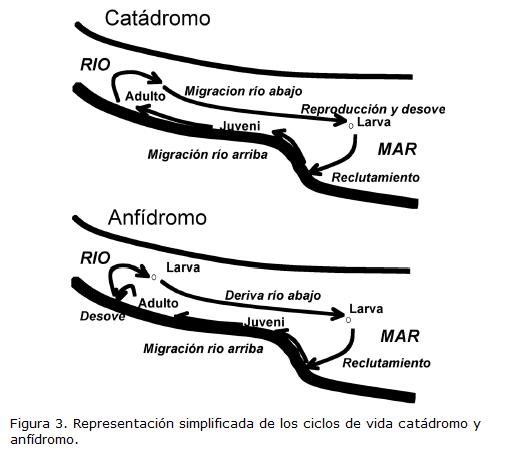
De acuerdo con McDowall (McDowall, 1988; McDowall, 1993a), un prominente ictiólogo neozelandés que ha dedicado su vida al estudio de los peces diádromos (Jellyman, 2002), existen tres variaciones de este tipo de estrategia de vida. El tipo anádromo es el más conocido porque lo presentan los salmónidos, los cuales después de nacer en las cabeceras de los ríos de zonas templadas, derivan por el río hasta llegar al mar, donde permanecerán hasta alcanzar la madurez sexual, para luego retornar a su río natal y reproducirse. El ejemplo clásico de pez catádromo es la anguila, la cual nace y se desarrolla en el mar, pero se desplaza a los estuarios y ríos hasta convertirse en adulto (Fig. 3). Posteriormente retorna al mar para reproducirse. El tipo anfídromo ha sido menos estudiado, a pesar de que la mayoría de peces de islas como Nueva Zelanda poseen este ciclo de vida (McDowall, 1988; McDowall, 1993a). En éstos, al igual que en los anádromos, los individuos nacen en el río y derivan hasta el mar, sin embargo regresan a los estuarios y suben por los ríos como juveniles pero esta migración es lenta y les puede tomar toda la vida, llegándose a convertir en adultos a lo largo de dicho viaje (Fig. 3). Por ejemplo, los caracoles anfídromos de la familia Neritidae pueden recorrer entre 1 y 50 m/día aguas arriba y cubrir >10 km en tres o cinco años (Blanco JF, datos sin publicar). La estrategia anfídroma predomina sobre la catádroma en las islas tropicales, en las cuales no se ha reportado hasta el momento la anadromía. Las primeras alusiones a la anfidromía aparecen en Myers, 1949, pero ha sido McDowall (McDowall, 1988; McDowall, 1993a; McDowall, 2004) quien ha abogado por el pleno reconocimiento de este tipo de estrategia. De hecho él y otros colegas (Gross et al., 1988) propusieron que una mayor productividad de las aguas dulceacuícolas en las islas tropicales y subtropicales que la de las aguas costeras podría explicar la prevalencia de los ciclos catádromos y anfídromos en ellas. El patrón inverso se observa en las zonas templadas más allá de los 30° de latitud en el hemisferio norte. Sin embargo, el mismo McDowall, co-proponente de la hipótesis de la productividad, ha sugerido recientemente (McDowall, 2008) que dicho mecanismo debe ser interpretado con cuidado ya que solo explica una fracción de la diversificación y dominancia de la estrategia anádroma en altas latitudes, principalmente la boreal. Llama especialmente la atención sobre el hecho de que en Nueva Zelanda, a pesar de estar ubicada en la zona templada austral, haya pocas especies anádromas y predominen las catádromas y anfídromas, o que en la Patagonia no haya ninguna especie anádroma. Por lo tanto, es necesario explorar hipótesis alternativas sobre el origen de la catadromía y la anfidromía.
La diadromía es, sin embargo, una estrategia aparentemente exitosa debido a su evolución convergente en repetidas ocasiones en varios linajes que ocupan las quebradas insulares. Por ejemplo, se ha documentado en gasterópodos, camarones y peces, lo que significa que apareció por primera vez en el Cámbrico (hace 550 millones de años) en los moluscos, luego en el Silúrico (hace 440 millones de años) en los peces y finalmente en los camarones en el Devónico (hace 410 millones de años). Aún dentro de los peces ha aparecido desde los Agnatha hasta los Gobioidei (McDowall, 1993b).
Con respecto al origen de la estrategia diádroma, coloquialmente se asume que es marino (discutido por McDowall, 2004), contrario al origen dulceacuícola planteado por Darwin. Los estudios recientes sugieren que la diadromía (McDowall, 1993b), y particularmente la anfidromía (McDowall, 2004), se han originado indistintamente de ancestros marinos y dulceacuícolas primarios. Lo que sí es consistente es que la diadromía es un carácter derivado ya que ningún grupo basal lo posee. Más aun, recientes estudios moleculares indican que existe un progresivo distanciamiento genético de los camarones anfídromos de varias especies de Atyidae desde el Indopacífico hacia el Caribe (Page et al., 2008) y por lo tanto ha existido una radiación antigua (Eoceno-Mioceno) dentro de la familia, y en consecuencia un ancestro anfídromo ha dado origen a un linaje completo de géneros anfídromos. Por otra parte, Page et al., 2008, también resaltan que aun las especies anfídromas podrían requerir de los -caminitos- (stepping stones) para dispersarse a través de cuencas oceánicas, debido a que su reproducción ocurre en los ríos y quebradas y no en el mar, contrario a lo que ocurre en las catádromas. Por esta razón, aunque las especies anfídromas tienen amplios ámbitos de distribución, no llegan a ser cosmopolitas. De acuerdo al planteamiento darwiniano de la dispersión ancestral, los camarones Atyidae tuvieron que dispersarse desde el Indopacífico hacia el Caribe -cuando las condiciones ambientales eran benignas-, es decir antes del cierre del istmo de Panamá.
LA DIADROMIA COMO FACTOR SELECTIVO EN LAS QUEBRADAS INSULARES
Darwin en el capítulo 12 de El origen planteó que:
No debemos olvidar que para tener un amplio rango de distribución no solo se necesita tener el poder de cruzar barreras si no que es más importante tener el poder de ser victorioso en la lucha por la vida con las especies foráneas.
Hasta el momento se ha demostrado que la gran capacidad de las especies con estrategias diádromas para cruzar la barrera oceánica e invadir aguas dulces en islas y costas lejanas. Pero también ellas pueden cruzar barreras como las cascadas ubicadas a lo largo de ríos y quebradas (Fièvet, 2000). Se ha publicado que algunos peces y camarones pueden trepar por las paredes de las cascadas o rodearlas saliéndose del cauce. Sin embargo, es claro que la mayoría de especies diádromas no pueden cruzar cascadas muy altas. Esto genera un mosaico de tramos: -aguas abajo- de las quebradas con presencia de especies diádromas y -aguas arriba- con ausencia de ellas. Esto tiene un efecto importante para las comunidades de insectos e invertebrados acuáticos y para los procesos ecosistémicos. Por ejemplo, algunos camarones (Atyidae y Xiphocariidae) son detritívoros y bioperturbadores muy activos que pueden no solamente contribuir al procesamiento de material vegetal y sedimentos orgánicos, sino también a ejercer una fuerte competencia sobre los insectos herbívoros, colectores y detritívoros (Pringle et al., 1999; Chong et al., 2000; Crowl et al., 2001). Por otra parte, algunos peces y camarones depredadores ejercen un control numérico sobre sus presas, pero también pueden ejercer una presión que selecciona caracteres morfológicos que posiblemente actúan como defensas (Covich et al., 2009). Estas presiones selectivas son aun más fuertes si se tiene en cuenta que algunas especies de camarones diádromos pueden vivir entre 5 y 13 años, lo cual es significativo entre los invertebrados acuáticos, tal como lo han establecido estudios de campo en Hong Kong y Puerto Rico (Mantel y Dudgeon, 2004; Cross et al., 2008).
LA ISLA GORGONA Y SU FAUNA LOTICA COMO MODELO DE ESTUDIO
Gorgona es la isla continental más grande del Pacífico colombiano y posee un número significativo de cursos de agua permanente (>25). Puede considerarse una isla pequeña (2,5 x 9 km) pero cercana al continente (a 35 y 52 km de los puntos más cercanos). Por esta razón ningún orden de Insecta se encuentra ausente. Sin embargo, la abundancia y riqueza son menores y la composición taxonómica es diferente a la del continente lo que resalta la naturaleza de la barrera oceánica. Por otro lado, los insectos coexisten con varias especies de peces, camarones y gasterópodos diádromos, los cuales son más abundantes. Es posible que esta fauna sea un relicto de una dispersión ancestral, ya que se comparten varias especies con el Caribe como el pez Agonostomus monticola (Mugilidae) y el camarón Potimirin glabrata (Atyidae). Otras especies son típicas del Pacífico oriental tropical pero pertenecen a géneros pantropicales (e.g. Macrobrachium, Atya, Neritina). Finalmente, existen un cangrejo (Hippolobocera gorgonae) y un pez (Trichomycterus gorgonae) primariamente dulceacuícolas endémicos. Estos hacen parte de géneros típicos de las quebradas de cabeceras andinas y por lo tanto apoyan la hipótesis de un puente de dispersión entre Gorgona y el continente. Se ha planteado que Gorgona está situada sobre la cuarta cordillera andina (actualmente sumergida) que se desprende desde Ecuador y que hizo parte del antiguo valle del río Patía durante la última glaciación (Kerr, 2005). Por lo tanto, la isla Gorgona ofrece una oportunidad única a nivel nacional para estudiar la ecología y biogeografía de los sistemas lóticos insulares. Recientemente se completó un compendio sobre el tema (Blanco et al., 2009).
LAS REPRESAS: UNA NUEVA PRESIÓN SELECTIVA PARA LA DIADROMIA
Los países tropicales están viviendo la edad dorada de la construcción de represas (Ramírez et al., 2008). Esto puede tener implicaciones para la ecología de los sistemas lóticos insulares y particularmente para la evolución de las especies diádromas. Se ha registrado bien en Puerto Rico y Nueva Zelanda que aguas arriba de las represas altas no existen especies de peces diádromos debido a la barrera física que éstas imponen para la migración (Joy y Death, 2001; Greathouse et al., 2006). Más aún, en ríos donde las cabeceras eran dominadas por especies diádromas, posteriormente a la construcción de las represas se han extinguido dichas especies y han sido reemplazadas por especies exóticas primariamente dulceacuícolas. Aunque todo apunta a una extinción generalizada de especies diádromas en las cabeceras de los ríos insulares y costeros posterior al represamiento, las represas son un cuello de botella que puede forzar nuevos eventos evolutivos. Recientemente se publicó que una especie de pez diádromo (Eleotridae: Gobiomorus dormitor) se estaba reproduciendo aguas arriba de una represa en Puerto Rico, sin necesidad de que sus larvas tuviesen que ir al mar para continuar con su desarrollo (Bacheler et al., 2004). Esto tiene unas implicaciones evolutivas importantes porque sugiere que al seleccionarse un genotipo completamente dulceacuícola, la población puede mantenerse en ausencia de la conexión con el mar. Sin embargo, esto no la libera de los riesgos que enfrentan las poblaciones pequeñas como la mortalidad catastrófica, las enfermedades y la endogamia, los cuales podrían experimentar con mayor probabilidad ahora aisladas en las cabeceras de cuencas particulares.
CONCLUSIONES
Darwin contribuyó al entendimiento de la ecología evolutiva dulceacuícola insular al abordar los modelos de dispersión de su fauna, particularmente de los insectos y peces. Él también vislumbró la diadromía como una novedad evolutiva en las islas oceánicas. Este ensayo resalta la importancia de la diadromía como control importante de procesos ecológicos y evolutivos en las quebradas insulares. Finalmente, las observaciones de Darwin y las investigaciones recientes demuestran que las quebradas de las islas oceánicas presentan entomo– e ictio-faunas dulceacuícolas poco abundantes y pobres en especies (deficientes en algunos grupos y con algunas rarezas) y por lo tanto pueden considerarse vacías. Sin embargo, la evolución convergente y recurrente de la estrategia diádroma las ha -llenado- con peces, camarones y gasterópodos que desarrollan su ciclo de vida entre el mar y las aguas dulces. La isla Gorgona en Colombia ofrece una oportunidad única para estudiar la ecología y biogeografía lótica insulares.
AGRADECIMIENTOS
El autor agradece a Eugenio Andrade y al comité organizador por la invitación para deponer en la Cátedra -Darwin 200 años-. Esta reflexión se ha nutrido de la investigación pasada y en curso del autor en Puerto Rico y Gorgona. Se agradece al comité de apoyo a la investigación de la Universidad de Antioquia por la financiación del proyecto -Gorgona Island Stream Bioassessment-. La investigación en Puerto Rico ha sido financiada desde 2000 por el Programa de investigaciones ecológicas a largo plazo en el bosque experimental de Luquillo, el Instituto internacional de dasonomía tropical del Servicio forestal de los Estados Unidos y la Universidad de Puerto Rico, Río Piedras. Los comentarios de dos evaluadores anónimos contribuyeron a mejorar el manuscrito.
BIBLIOGRAFÍA
BACHELER NM, NEAL JW, NOBLE RL. Reproduction of a Landlocked Diadromous Fish Population: Bigmouth Sleepers Gobiomorus dormitor in a Reservoir in Puerto Rico. Caribb J Sci. 2004;40:223-231.
[ Links ]BASS D. A Comparison of Freshwater Macroinvertebrate Communities on Small Caribbean Islands. Bioscience. 2003;53:1094-1100.
[ Links ]BENSTEAD JP, MARCH JG, PRINGLE CM, EWEL KC, SHORT JW. Biodiversity and Ecosystem Function in Species-Poor Communities: Community Structure and Leaf Litter Breakdown in a Pacific Island Stream. J North Am Benthol Soc. 2009;28:454-465.
[ Links ]BLANCO JF, RAMIREZ A, SCATENA FN. The Streams of Gorgona Island National Natural Park within the Global Context: An Introduction to the Special Issue. Actu Biol. 2009;31(91):105-110.
[ Links ]BLANCO JF, SCATENA FN. Floods, habitat hydraulics and upstream migration of Neritina virginea (Gastropoda: Neritidae) in Northeastern Puerto Rico. Caribb J Sci. 2005;41:55-74.
[ Links ]BRASHER AMD. Impacts of Human Disturbances on Biotic Communities in Hawaiian Streams. Bioscience. 2003;53:1052-1060.
[ Links ]BROWN JH, LOMOLINO MV. Biogeography. 2 ed. Sunderland (MA): Sinauer; 1998.
[ Links ]CHONG CT, LARNED ST, COVICH AP, KINZIE III RA. Species Interactions between Estuarine Detritivores: Inhibition or Facilitation? Hydrobiologia. 2000;434:11-16.
[ Links ]COVICH AP. Dispersal-limited Biodiversity of Tropical Insular Streams. Pol J Ecol. 2006;54:523-547.
[ Links ]COVICH AP, CROWL TA, HEIN CL, TOWNSEND MJ, MCDOWELL WH. Predator-Prey Interactions in River Networks: Comparing Shrimp Spatial Refugia in Two Drainage Basins. Freshw Biol. 2009;54:450-465.
[ Links ]CRAIG DA. Geomorphology, Development of Running Water Habitats, and Evolution of Black Flies on Polynesian Islands. Bioscience. 2003;53:1079-1093.
[ Links ]CROSS WF, COVIC AP, CROWL TA, BENSTEAD JP, RAMIREZ A. Secondary Production, Longevity and Resource Consumption Rates of Freshwater Shrimps in Two Tropical Streams with Contrasting Geomorphology and Food Web Structure. Freshw Biolo. 2008;53:2504-2519.
[ Links ]CROWL TA, MCDOWELL WH, COVICH AP, JOHNSON SL. Freshwater Shrimp Effects on Detrital Processing and Localized Nutrient Dynamics in a Montane, Tropical Rain Forest Stream. Ecology. 2001;82:775-783.
DARWIN CR. Journal and Remarks. 1832-1836. London: Henry Colburn; 1839. Disponible en: URL: http://darwin-online.org.uk/contents.html; 1839 [ Links ] [ Links ]DARWIN CR. The Zoology of the Voyage of H.M.S. Beagle. London: Henry Colburn; 1838-1843. Disponible en: URL: http://darwin-online.org.uk/contents.html; 1838-1843
[ Links ]DARWIN CR. The Origin of the Species. London: Bantam Classic Book; 1859.
[ Links ]FIÈVET E. Passage Facilities for Diadromous Freshwater Shrimps (Decapoda: Caridea) in the Bananier River, Guadeloupe, West Indies. Regul River. 2000;16:101-112.
[ Links ]GILLESPIE RG, RODERICK GK. Arthropods on Islands: Colonization, Speciation, and Conservation. Annu Rev Entomol. 2002;47:595-632.
[ Links ]GREATHOUSE EA, PRINGLE CM, HOLMQUIST JG. Conservation and Management of Migratory Fauna: Dams in Tropical Streams of Puerto Rico. Aquat Conserv. 2006;16:695-712.
[ Links ]GROSS MR, COLEMAN RM, MCDOWALL RM. Aquatic Productivity and the Evolution of Diadromous Fish Migration. Science. 1988;239:1291-1293.
[ Links ]JELLYMAN D. Bob McDowall-His Contribution to New Zealand s freshwater fish. N Z J Mar Freshwater Res. 2002;36:1-12.
[ Links ]JOY MK, DEATH RG. Control of Freshwater Fish and Crayfish Community Structure in Taranaki, New Zealand: Dams, Diadromy or Habitat Structure? Freshw Biol. 2001;46:417-429.
[ Links ]KERR AC. La Isla de Gorgona, Colombia: A Petrological Enigma. Lithos. 2005;84:77-101.
[ Links ]MACARTHUR RH, WILSON EO. The Theory of Island Biogeography. Princeton (NJ): Princeton University Press; 1967.
[ Links ]MCDOWALL RM. Diadromy in fishes: migrations between freshwaters and marine environments. London: Croom Helm; 1988.
[ Links ]MCDOWALL RM. Implications of Diadromy for the Structuring and Modelling of Riverine Fish Communities in New Zealand. N Z J Mar Freshwater Res. 1993a;27:453-462.
[ Links ]MCDOWALL RM. A Recent Marine Ancestry for Diadromous Fishes? Sometimes yes, but mostly no. Environ Biol Fishes. 1993b;37:329-335.
[ Links ]MCDOWALL RM. Ancestry and Amphidromy in Island Freshwater Fish Faunas. Fish Fish. 2004;5:75-85.
[ Links ]MCDOWALL R. Why are so Many Boreal Freshwater Fishes Anadromous? Confronting -Conventional Wisdom-. Fish Fish. 2008;9:208-213.
[ Links ]MANTEL SK, DUDGEON D. Growth and Production of a Tropical Predatory Shrimp, Macrobrachium hainanense (Palaemonidae), in two Hong Kong Streams. Freshw Biol. 2004;49:1320-1336.
[ Links ]MARCH JG, BENSTEAD JP, PRINGLE CM, LUCKYMIS M.A. Benthic Community Structure and Invertebrate Drift in a Pacific Island Stream, Kosrae, Micronesia. Biotropica. 2003;35:125-130.
[ Links ]MYERS GS. Usage of anadromous, catadromous and allied terms for migratory fishes. Copeia. 1949;1949:89-97.
[ Links ]PAGE TJ, COOK BD, RINTELEN T, RINTELEN K, HUGHES JM. Evolutionary Relationships of Atyid Shrimps Imply both Ancient Caribbean Radiations and Common Marine Dispersal. J North Am Benthol Soc. 2008;27:68-83.
[ Links ]PRINGLE CM, HEMPHILL N, MCDOWELL W, BEDNAREK A, MARCH JG. Linking Species and Ecosystems: Different Biotic Assemblages Cause Instream Differences in Organic Matter. Ecology. 1999;80:1860-1872.
[ Links ]RAMÍREZ A, PRINGLE CM, WANTZEN KM. Tropical Stream Conservation. En: Dudgeon W, editor. Tropical stream ecology. London: Academic Press (Aquatic Ecology Series); 2008. p. 285-304.
[ Links ]SMITH GC, COVICH AP, BRASHER AMD. An Ecological Perspective on the Biodiversity of Tropical Island Streams. Bioscience. 2003;53:1048-1051.
[ Links ]VINSON MR, HAWKINS CP. Broad-scale geographical patterns in local stream insect genera richness. Ecography. 2003;26:752-767.
[ Links ]