








Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAvances en Psicología Latinoamericana
Print version ISSN 1794-4724On-line version ISSN 2145-4515
Av. Psicol. Latinoam. vol.28 no.1 Bogotá Jan./June 2010
Esquizofrenia, genética, epigênesis, ambiente: uma revisão sistemática das hipóteses etiológicas unificadas e do perfil genética; e um novo algoritmo para o tratamento dos achados principais*
Schizophrenia, genetics, epigenesis, environment: a systematic review of unified etiological hypotheses and genetic profile; and a new algorithm to cope with the main findings
Álvaro Machado Dias**
E Avelino Luiz Rodrigues***
* Agradecimentos: Agradecemos à equipe de Hervé Rostaing da Universidade Paul Cézanne Aix-Marseille-III e especialmente a Bruno Mannina por ter-nos dado o devido suporte técnico, bem como por nos ter acompanhado em discussões técnicas sobre a implementação das funções requeridas pela presente revisão. Paralelamente, gostaríamos de destacar nosso agradecimento aos criadores das ferramentas gratuitas Ali Baba (http://alibaba.informatik.hu-berlin.de/), Arrowsmith (http://arrowsmith.psych.uic.edu/arrowsmith_uic/index.html) e Medie (http://www-tsujii.is.s.u-tokyo.ac.jp/medie/) que nos ajudaram a refinar a seleção bibliográfica, utilizada ao longo da produção deste paper. Uma versão comercial do Matheo-analizer (sem a função semântica aludida) pode ser encontrada em: http://www.matheo-analyzer.com/.
** Álvaro Machado Dias PhD, Departamento de Psiquiatria, Faculdade de Medicina da Universidade de São Paulo, Rua Ovídio Pires Campos s/n., São Paulo, Brasil. Tel.: 0055-11-3091-1901, mail: alvaromd@usp.br.
*** Médico psiquiatra, Coordenador do Laboratório de Pesquisa Sujeito e Corpo do IP-USP e Professor Doutor do Dep. de Psicologia Clínica da Universidade de São Paulo (PSC-USP). Av. Prof. Mello Moraes, 1721 Caixa Postal 66261 Cidade Universitária, São Paulo, Brasil. Tel.: 0055-11-3091-1901, mail: avelinoluizr@usp.br.
Recepción: 6 de abril de 2009
Aceptación: 23 de julio de 2009
Abstract
Context: schizophrenia is a highly complex syndrome, related to genes, and to non-genetic risk factors. Famous epidemiological studies reported its presence among all cultures and geographic regions. In that sense, Unified Etiological hypothesis face the challenge to both present experimental data, and to show that the findings may cope with the syndromes universal profile. Objectives: systematically review the most prominent Unified Etiological hypothesis, as much as the semantic distribution of genetic findings (under up to date data mining techniques), and propose a new model, based on the dynamic effects of epigenics over genetic activation in both neurodevelopment and early adulthood. Results: in general, Unified Etiological Hypothesis contradict the main genetic findings (which suggest that schizophrenias genes are mostly associated with neurotransmitter profiles, like D-1 and the Glutamate-NMDA cascade); also in general, genetic findings are spread all over the genome (as we reveal with a topological map of the 3519 studies on the matter). The key for this conundrum may be represented by the association between the perspective that each polymorphism associated with schizophrenia represents a statistical risk factor (e.g. increasing the risk of developmental instability) while epigenetic molecular cascades and environmental factors considerably influence this picture, affecting genetic activation within critical periods.
Keywords: schizophrenia, etiology, brain.
Resumen
Contexto: la esquizofrenia es un síndrome complejo relacionado con genes y factores de riesgo no genéticos. Estudios epidemiológicos reconocidos reportan su presencia en todas las culturas y regiones geográficas. En este sentido, las Hipótesis Etiológicas Unificadas enfrentan simultáneamente el desafío de presentar los datos experimentales y demostrar que éstos dan cuenta del perfil universal del síndrome. Objetivos: revisar sistemáticamente las más prominentes Hipótesis Etiológicas Unificadas, así como la distribución semántica de los hallazgos genéticos (mediante técnicas de minería de datos) y proponer un nuevo modelo, basado en los efectos dinámicos de carácter epigenético sobre la activación genética en el neurodesarrollo y la pubertad. Resultados: de manera general, las Hipótesis Etiológicas Unificadas contradicen los principales hallazgos genéticos (que sugieren que la esquizofrenia está asociada al perfil de neurotransmisores como D-1 y la cascada Glutamato-NMDA); también, por regla general, los hallazgos genéticos se encuentran esparcidos por todo el genoma (tal como revelamos en un mapa topológico de los 3519 estudios en el asunto). La clave para este estado complejo de cosas puede estar representada por la asociación entre la perspectiva de que cada polimorfismo asociado a la esquizofrenia representa un factor de riesgo estadístico (es decir, aumentando el riesgo de inestabilidades del desarrollo), mientras que las cascadas moleculares de carácter epigenético y los factores de riesgo ambientales permanecen actuantes mediante la activación de genes en períodos críticos.
Palabras clave: esquizofrenia, etiología, cerebro.
Introdução
Tal como em muitas outras décadas, avanços recentes nos permitem asseverar que uma nova concepção sobre a esquizofrenia vem emergindo consistentemente. Dito isto, destaca-se também que, tal como muitas outras, nesta primeira década do século XXI as perguntas permanecem superando em muito as respostas e as novas concepções não resolvem assuntos complexos como bases neurais e tratamento.
A primeira vista, seria de se considerar que, em domínio etiológico, a perspectiva não fosse diferente; não obstante ela é – não em sentindo resolutivo, mas tanto pelo contrário, por força do que se convencionou chamar paradoxo etiológico da esquizofrenia, uma prerrogativa de cunho conceitual, relativa à prevalência da síndrome na atualidade a despeito de seu caráter desadaptativo, que torna a problemática etiológica particularmente delicada.
Considerando este panorama, o objetivo do presente artigo é: 1. Revisar sistematicamente a literatura em etiologia da esquizofrenia, através de princípios qualitativos (apresentação das hipóteses principais e, subseqüentemente, apresentação de evidências; 2. Revisar a literatura, a partir de um método em franco desenvolvimento: a extração de padrões de data mining (ver: métodos); 3. Introduzir um modelo etiológico consistente desde os dados anteriormente alinhavados.
Métodos
O presente artigo utiliza-se de dois métodos para tingir os resultados almejados: revisão manual e levantamento de evidências acerca dos tópicos de interesse; análise de correlações em ambiente de softwares voltados para o reconhecimento de padrões em sistemas extensivos de múltiplas variáveis (genomas, sistemas numéricos). Isto é, consideramos todas as referências indexadas na problemática abordada, para dela extrair o perfil quantitativo mais fidedigno possível.
Quanto a isto, destacamos a oportunidade concedida pelo grupo de Hervé Rostaing da Universidade Paul Cézanne Aix-Marseille-III de utilizar o software Matheo-analyzer 1.2.3 (também conhecido como Beta-2) com funções semânticas enriquecidas – e ainda não disponível no mercado.
Revisão sistemática da literatura em etiologia da esquizofrenia
Paradoxo etiológico
Ainda que o conceito de esquizofrenia seja recente e que a nosologia sistemática tenha cerca de um século de idade (ainda como dementia praecox por Kraepelin e então com Jaspers e Bleuler), é de se considerar que relatos de loucura com perfil próximo ao que convencionamos denominar esquizofrenia, já ocorressem durante a antiguida de (Pliskas, 2004; Wilson, 1981). Paralelamente, destaca-se que os portadores da síndrome geram em média cerca de metade do número de filhos de seus pares não afetados (Bassett, Bury, Hodgkinson, & Honer, 1996). Não obstante, a prevalência de esquizofrenia se mantém na população mundial em considerável estabilidade, girando em torno de 0.01, sendo que grandes estudos epidemiológicos, tal como os diversos estudos multicêntricos da organização mundial da saúde (WHO), sugerem que a síndrome ocorra em todas as culturas e áreas geográficas, através de taxas bastante semelhantes (Jablensky & Sartorius, 1988).
Como é de se considerar, esta conjuntura é paradoxal, na medida em que a desordem leva seus portadores a produzir menos dependentes e não parece ser fruto de condições recentes (o que de maneira alguma exclui a importância de variáveis culturais para a precipitação dos sintomas e prognóstico). Neste sentido, o paradoxo etiológico se traduz pela premência em se explicar porque a síndrome não foi eliminada da população.
A epistemologia das hipóteses etiológicas de tipo teórico
Ao contrário do que ocorre com a maioria das doenças mentais, tais como a síndrome de Down, muitas das hipóteses etiológicas da esquizofrenia freqüentemente preconizam amplas refl exões auxiliares à apresentação dos dados empíricos que lhes suportariam, isto é, grande parte destas hipóteses não se limita a relacionar achados clínicos a achados biológicos subjacentes, mas antes propõe uma maneira de se interpretar os mesmos à luz de paradigmas definidos pelo autor em questão.
Isto se explica porque, ainda que cada nova hipótese se escore em achados que melhor lhe convenham para sugerir um caminho à compreensão do fator etiológico, dificilmente alguma é capaz de passar pelo teste da história enquanto se faça insuficiente à luz do paradoxo etiológico, sendo esta a sua derradeira importância para o campo como um todo.
Por exemplo, consideremos uma hipótese que se proponha como Hipótese Etiológica Unificada da Esquizofrenia, associando a síndrome a uma mutação consistentemente encontrada no genoma dos portadores (i.e. COMT, Neurogulin, DISC-1, etc.); uma maneira de se asseverar o potencial da mesma enquanto tal (Hipótese Unificada) dá-se pela sondagem da penetrância da mutação ao longo de diversas populações. Dada a prevalência global da esquizofrenia, a demonstração de que a mutação em questão está restrita a determinadas populações, limita sua generalidade e, portanto, sua representatividade. Dir-se-ia: tal hipótese é incapaz de superar as barreiras do paradoxo etiológico, já que não revela a distribuição de 1% ao longo de toda a população e, assim, não explica a permanência do traço. É precisamente esta conjuntura que revela porque as hipóteses freqüentemente aludem a variáveis que não podem ser diretamente sondadas (ou não são falseáveis)
Tendo em vista as características centrais das mais conhecidas e infl uentes hipóteses etiológicas de inspiração teórica, dir-se-ia que existem três tipos clássicos potencialmente compatíveis com a dissolução do paradoxo etiológico da esquizofrenia. As hipóteses que apontam para um ou mais aspectos funcionais subjacentes à síndrome (doravante denominadas funcionais em sentido forte); as hipóteses que apontam para a possibilidade de ter a síndrome surgido em conjunção com algum traço adaptattivo, sem carregue em seu bojo qualquer característica vantajosa (doravante denominadas funcionais em sentido fraco); e as hipóteses que sugerem que não são os genes especificamente associados ao pedigree dos portadores que levam à expressão sindrômica, mas antes cadeias de eventos irredutíveis ao DNA, e tanto mais ligados à transcrição e expressão dos genes (doravante denominadas epigênicas).
Hipóteses funcionais em sentido forte mais conhecidas
Considerando as hipóteses funcionais em sentido forte em ordem cronológica, dir-se-ia que a primeira a se tornar amplamente conhecida e debatida nos meios científicos foi a de Huxley e colaboradores (1964) (subseqüentemente desenvolvido por Erlenmeyer-Kimling (1968), que sugeriu que a prevalência de esquizofrenia se relacione a um aumento na taxa de reprodução dos descendentes diretos dos portadores da síndrome, em função de um único gene.
Outra hipótese bastante comentada é de Oken, que percebeu a ocorrência de baixa taxa de artrite reumatóide entre os portadores da síndrome (0.66% em comparação à taxa média da população em geral de 1%), daí extraindo uma medida de sua potencial funcionalidade (Oken e Schulzer, 1999). Um aspecto anedótico desta (mas digno de nota) é o de que se trata de uma das únicas hipóteses desta natureza construídas a partir de uma meta-análise: foi justamente de uma revisão de literatura, reunindo 14 estudos em que 12 reportavam negativamente a presença de artrite reumatóide nos portadores de esquizofrenia, que os autores construíram a hipótese.
Uma terceira hipótese notável é a de Brown, denominada hipótese infecciosa da esquizofrenia, a qual relaciona a síndrome à maior resistência a infecções e demonstra, no âmbito da literatura em genética das infecções e modelos animais de genética da esquizofrenia, que as regiões de interesse genômico são próximas (James S. Brown Jr, 2003) e tornam sugestiva a possibilidade que estar a esquizofrenia relacionada a um polimorfismo conferindo vantagem ou sobrevivência, especialmente durante epidemias (p. 109).
Hipóteses funcionais em sentido fraco mais Conhecidas
Entre as hipóteses funcionais em sentido fraco mais conhecidas, destaca-se inicialmente a hipótese da esquizofrenia como fruto da seleção sexual (Shaner, Miller e Mintz, 2004), que sugere que a esquizofrenia represente um extremo desadaptativo da habilidade sexualmente selecionada do cortejo (104), isto é, os polimorfismos que contribuem para a etiologia da esquizofrenia seriam os mesmos que, em condições ideais, levam ao desenvolvimento de habilidades (verbais) selecionadas sexualmente.
É de se considerar que esta hipótese representa uma derivação da hipótese funcional em sentido fraco mais conhecida, denominada hipótese da hemisferização assimétrica, de Timothy Crow, (2006; 2008), cujo mote é o de que a síndrome represente um preço a se pagar pela evolução do cérebro humano, isto é, uma vicissitude populacional indesejável da emergência de um traço (a arquitetura cognitiva do Homo Sapiens sapiens) vantajoso.
Sugere-se com esta hipótese, que a síndrome emerja no contexto do processo de migração neural para a consolidação da assimetria entre os hemisférios cerebrais. Como é de se considerar, esta assimetria favorece (na maioria da população) o hemisfério esquerdo, onde se concentram as áreas de linguagem. Desde esta premissa, o autor enfatiza que perdas de assimetria inter-hemisférica se revertem em menores capacidades cognitivas e outros sintomas associados direta ou indiretamente à síndrome.
Em termos neuroanatômicos, a hipótese se fia na menor assimetria entre os hemisférios do cérebro na população clínica – e em consonância ao desempenho cognitivo diminuído em testes de atenção e memória de trabalho e do que média da população humana (Berlim, Mattevi, Belmonte-de-Abreu, & Crow, 2003). Crow concebe assim a esquizofrenia como uma encefalopatia genética dos circuitos fronto-temporais relacionados à linguagem, sendo a manutenção dos genes relacionados à esquizofrenia no pool genético da espécie relativa ao pouco que tempo transcorrido desde o surgimento da linguagem, o que por sua vez não teria permitido a eliminação do traço.
Inconsistências das hipóteses funcionais
O modelo de Huxley e colaboradores (1964) fazse problemático no sentido em que se sabe que a síndrome não é fruto de um único gene, bem como as taxas de reprodução individuais estão diminuídas em 50%, o que torna o custo/benefício de um eventual aumento na taxa de reprodução de descendentes um fator de compensação desfavorável (Bassett, et al., 1996).
O modelo de Oken, que relaciona baixa taxa de artrite reumatóide ao risco de esquizofrenia é contraditório a dados mais atuais que sugerem que as doenças auto-imunes em geral são mais presentes em famílias de portadores da síndrome e assim justamente representam fator de risco (no sentido prospectivo de risco). Um estudo com cerca de 7700 portadores da síndrome concluiu que a posse de alguma doença auto-imune aumenta a chance de se manifestar esquizofrenia em 45% (Eaton, et al., 2006).
O modelo infeccioso da esquizofrenia é igualmente pouco plausível conquanto os principais genes mais fortemente relacionados à síndrome (que consideraremos abaixo) não estão elencados entre aqueles apontados por Brown (2003).
A hipótese da seleção sexual é a princípio pertinente, já que as taxas de reprodução dos portadores da síndrome de fato são diminuídas, enquanto a mesma costuma se manifestar no final da puberdade, período em que a vida sexual assume o seu rumo pleno. Não obstante, é de se ter em vista que, rigorosamente falando, este tipo de argumento só faz sentido para desordens que afetem exclusiva ou majoritariamente homens e que, privilegiadamente, relacionem-se à transmissão através de cromossomos sexuais, o que parece não ser o caso para a esquizofrenia, como veremos abaixo. Um estudo da organização mundial de saúde (WHO) em 10 países sugere que as taxas de esquizofrenia sejam homogêneas entre os sexos (Jablensky, et al., 1992). Outro estudo, contemplando cerca de 10 milhões de pacientes de fato sugere maior prevalência em homens, mas não em nível suficiente para que se faça plausível a afirmação de que a síndrome esteja relacionada a um traço cuja origem biológica é claramente relacionada à ação masculina em primatas e outros mamíferos.
Neste mesmo sentido, a hipótese da hemisferização assimétrica permite previsões e interpretações consistentes. Por exemplo, sob a vigência do modelo seria de se esperar que crianças ambidestras apresentassem predisposição maior do que destras para manifestação de esquizofrenia, após a adolescência. Em um estudo cruzando os dados de 16980 crianças que participaram de um programa inglês de estudo do desenvolvimento (NCDS) e dados de admissão dos sujeitos em hospitais psiquiátricos, (T. J. Crow, Done, & Sacker, 1996) encontraram que crianças pré-esquizofrênicas tinham maior tendência a serem ambidestras, sinalizando para um atraso nas manifestações de hemisferização assimétrica.
Não obstante, sabe-se atualmente que os efeitos cerebrais da esquizofrenia estão espalhados por todo o cérebro, posto que se relacionam fortemente a cascatas neurotransmissoras bastante difusas (i.e. glutamato-NMDA ; ver: Krystal, et al., 2003) e à estrutura da substância branca, a qual define as networks, simultaneamente em níveis local e global e se encontra afetada de maneira geral na esquizofrenia (Andreone, et al., 2007; Ashtari, et al., 2007; Buchsbaum, et al., 1998; Davis, et al., 2003; Lim, 2007; McGlashan e Hoffman, 2000; Schlosser, et al., 2007; Zanetti, et al., 2008; Zhou, et al., 2008). Faz-se ainda discutido se estas perdas necessariamente acompanham baixo desempenho cognitivo, vicissitude contradicta por algumas evidências (Mayringer e Wimmer, 2002) . Por fim, é de se ter em vista a própria relativização da associação entre perda d assimetria inter-hemisférica e esquizofrenia. Por exemplo: comparando gêmeos monozigóticos (n=10) concordantes e discordantes para a síndrome, um estudo conhecido (Bartley, Jones, Torrey, Zigun, e Weinberger, 1993) não encontrou diferenças no grau de assimetria entre os discordantes. O que esta conjuntura sugere é que, não sendo o caso de se negar a associação entre perdas de assimetria e probabilidade aumentada de se desenvolver esquizofrenia, é de se relativizar o elo causal, bem como o papel deste tipo de achado no âmbito das Hipóteses Etiológicas Unificadas da Esquizofrenia.
Revisão Quantitativa Dos Principais Achados Em Genética Da Esquizofrenia
Nesta seção apresentaremos resultados de um estudo de revisão quantitativa e sistemática da totalidade dos estudos publicados em genética da esquizofrenia (indexados no PubMed – ou – ISI), desenvolvido a partir da exploração de recursos em software para a análise de padrões complexos. A literatura contempla 3519 publicações no tema, indexadas nas bases de dados definidas acima. Destas se destaca que os genes mais comumente associados à esquizofrenia são: Neuregulin-1 (NRG-1), COMT e DISC-1. O primeiro está presente em 0.056 dos estudos; o COMT está presente em 0.080 dos estudos e DISC-1 está presente em 0.010 dos estudos, o que refl ete a pequena concordância do campo em torno dos achados.
O NRG-1 está ligado à atividade glutamatérgica e à estrutura da substância branca (Konrad e Winterer, 2008; Roy, et al., 2007), o COMT (através do alelo Val-158) relaciona-se à esquizofrenia através de cascatas para a inativação de catecolaminas (neste caso, com especial ênfase na dopamina; para uma meta-análise, ver: Fan, et al., 2005) e o DISC-1, com o neurodesenvolvimento, através de importantes cadeias proteicas para a formação do sistema nervoso central (para um editorial da BMJ sobre o gene, ver: Sawa e Kamiya, 2003).
Como é de se considerar, nenhum deles – assim como nenhum dos outros achados genéticos importantes da atualidade – pode ser associado aos argumentos anteriormente apresentados pelas Hipóteses Etiológicas Unificadas de cunho teórico. Isto não quer dizer que, por definição, devam ser descartadas, mas sugere que talvez careçam de reformulações ou ainda, que não versem exatamente sobre a esquizofrenia, mas antes sobre determinadas manifestações presentes em quadros sindrômicos e psicoses em geral, sem que possam ascender ao papel de explicação. Em outras palavras, talvez seja o caso de se considerar se as hipóteses não dizem respeito a um estado de coisas que se faz atingido em diversas situações de disfunção operacional do sistema nervoso com determinadas características, as quais se fazem presentes na esquizofrenia, mas nem por isto devem ser tomadas como a esquizofrenia.
De maneira geral, os 3519 estudos em genética da esquizofrenia obedecem à distribuição temática abaixo (termos em inglês refl etem a própria literatura), a qual nos revela os fatores associados, tanto em termos de loci, quanto variáveis endógenas primariamente afetadas (alvos) e comorbidades (Tabela 1).
Tabela 1. Distribuição dos temas em genética da esquizofrenia, nota-se a heterogeneidade das publicações
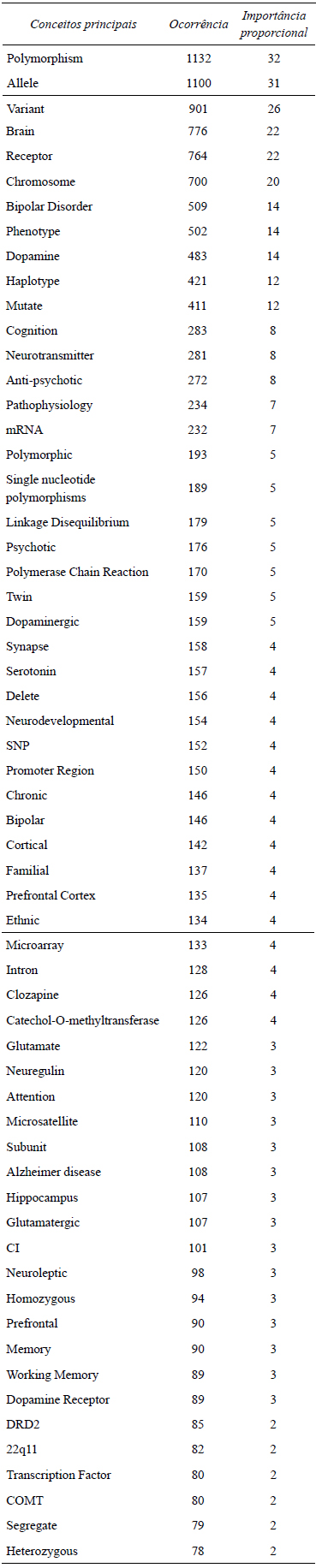
Note-e que o cromossomo mais relatado na literatura é o 22q11, onde se encontra o NRG-1. As habilidades cognitivas mais referidas como alvos dos polimorfismos associados à esquizofrenia são: memória e atenção. As áreas cerebrais mais representativas destas infl uências são: córtex pré-frontal e córtex temporal/hipocampo, enquanto a dopamina possui proeminência entre os neurotransmissores. Distúrbio bipolar desponta como proeminente comorbidade, sob os mesmos riscos genéticos. Por fim, como é de se notar, a heterogeneidade é a regra, não havendo um fator comum aos estudos. Disto decorre que os mesmos, organizam-se em função de seus achados (ou hipóteses genéticas) de acordo com o mapa topológico em a Figura 1.
Figura 1. Mapa de análise semântica da distribuição dos estudos de genética da esquizofrenia
Como se faz evidente do mapa, alguns achados são consistentemente replicados e criam clusters (principalmente os polimorfismos Neuregulin-1 e COMT), mas muitos outros se distribuem de maneira relativamente isolada em sub-populações específicas, ou mesmo não são replicados por grupos independentes de pesquisadores.
Em uma pesquisa na base de dados genômicos GeneCards encontramos 937 achados genéticos relacionados à esquizofrenia, e 17 Locus Genéticos alvos (BPP ERDA1 ETM1 NA NT3 SCZD1 SCZD10 SCZD11 SCZD12 SCZD13 SCZD2 SCZD3 SCZD4 SCZD5 SCZD6 SCZD7 SCZD8). A eles, podem-se acrescentar os resultados de um prestigioso estudo multicêntrico europeu de 2008, que testou a relação entre esquizofrenia e catorze dos principais nucleotídeos únicos associados pela literatura (RGS4, DISC1, DTNBP1, STX7, TAAR6, PPP3CC, NRG1, DRD2, HTR2A, DAOA, AKT1, CHRNA7, COMT e ARVCF), não encontrando relação significativa para nenhum deles, o que per se relativiza a atratividade de Hipóteses Etiológicas Unificadas em geral (Sanders, et al., 2008).
O caráter estratégico das hipóteses epigênicas
Como pudemos notar, o caráter heterogêneo dos achados e estudos em genética da esquizofrenia é a regra. Por estas razões, acreditamos que o futuro das hipóteses etiológicas da esquizofrenia alinha-se à proposição de desenvolvimentos contemplando a importãncia decisiva dos fatores epigênicos.
O conceito de epigênese é relativo aos mecanismos que entram em ação durante o processo de expressão de genes. A única hipótese epigênica sistematicamente desenvolvida na literatura denominase hipótese da instabilidade do desenvolvimento (DI), de Yeo e colaboradores. De acordo com esta, a esquizofrenia seria fruto de alterações no padrão de desenvolvimento ideal de um organismo, as quais seriam estatisticamente favorecidas pelos diversos fatores genéticos relacionados à esquizofrenia (Yeo, Gangestad, Edgar, e Thoma, 1999). Não obstante, a hipótese não define um papel para agentes não-genéticos, mas tão somente se diferencia das hipóteses anteriores pela relativização da importância de qualquer gene específico para a etiologia da síndrome em prol do efeito estatístico de cada um deles.
Para além dela, é de se notar os riscos aumentados relacionados à idade dos pais (Cheng, Ko, Chen e Ng, 2008), estacão climática de nascimento, devido às chances diversas de doenças e infecções durante a gravidez (para detalhes, ver: Bembenek, 2006; Bersani, et al., 2006; Cheng, et al., 2008; Gallagher, Jones, McFalls e Pisa, 2007; Jordaan, et al., 2006; Kallel, et al., 2007; Messias, et al., 2006), gripe e outras infecções viróticas da mãe durante a gestação (Ellman, Yolken, Buka, Torrey e Cannon, 2009; Yudofsky, 2009), infecção virótica perinatal (Fruntes e Limosin, 2008), estresse em períodos críticos (Rusch, et al., 2009) e muitos outros fatores não-genéticos.
Em função destes, bem como assumindo para si mesmos o fato de que a prática clínica com pacientes diagnosticados com esquizofrenia revela uma consistente relação entre manifestação, prognóstico e realidade psicológica (onde o estresse emerge como variável crítica), nesta derradeira seção propomos apresentar um algoritmo sistematizando o que denominamos hipótese epigênica estendida, sob a qual se assume que a pathway sindrômica não é gene/desenvolvimento neural/ alterações hormonais-realidade psicológica/manifestação, mas sim uma em que o conceito de gene como depositário da hereditariedade é subvertido em função da perspectiva (tanto mais atual) de que uma série de fatores infl uencia dinamicamente sua ligação/desligamento, tal como no algoritmo em a Figura 2.
Como pretendemos ilustrar, no modelo epigênico, a perspectiva é a de que os genes não sejam o derradeiro caminho à compreensão do processo de adoecimento, não por não serem crucial à etiologia, mas sim por estarem dinamicamente mediados pelos fatores epigênicos, por sua vez mediados pelos fatores ambientais (no sentido de extrínseco à constituição do organismo), dentro de uma chave em que as alterações hormonais da adolescência, amplamente infl uenciada pela ligação de genes, infl uenciam a derradeira etapa no processo de maturação neurológica, que no caso da esquizofrenia sabe-se estar pontualmente relacionada à perda de conformação conectiva cortical, eventualmente levada a cabo devido ao aumento desproporcional da poda neural (Hoffman e Dobscha, 1989), o que por sua vez naturalmente vai depender da ativação de genes, desde cascatas bioquímicas intermediadas por processos epigênicos e agentes ambientais extrínsecos. Como também é de se considerar, sob o nosso modelo pode-se falar em caso de esquizofrenia que não aconteceu, ou seja, pode-se conceber uma série de condições subclínicas que deixaram de evoluir à esquizofrenia por força da ausência de variáveis críticas.
Discussão e conclusão
Em nosso estudo revelamos (através de revisão sistemática) que as principais hipóteses etiológicas de inspiração funcional (baseadas na manutenção dos genes da esquizofrenia no pool genético da espécie em função de alguma vantagem ou associação contingente no ambiente adaptativo da espécie) não se sustentam. Mormente, apresentamos como contrapartida a perspectiva epigênica, cujo precursor é Yeo.
Dito isto, destacamos um algoritmo que transcende as posições de Yeo, na medida em que dinamiza e retroalimenta os efeitos ambientais-epigênicos sobre as cascatas bioquímicas que levam à esquizofrenia. Sob este mote, entende-se porque existem fenômenos como a esquizotipia e outras condições limítrofes não indentificadas e possivelmente relacionadas à presença de fatores de risco genéticos, em ambiente favorável à sua não-expressão.
O nosso modelo rechaça a idéia de que o ambiente em si possa produzir esquizofrenia - já que a cascata de eventos etiológicos depende da presença (e derradeira ação) de genes – ao mesmo tempo em que concede amplo potencial de destaque ás variáveis ambientais e psicológicas, as quais precisamente representam vias para a ativação de genes em diversos momentos críticos à emergência sindrômica. O que, diga-se de passagem, está alinhado à vasta maioria dos estudos atuais no tema.
Com estas concepções em vista é de se compreender que a própria necessidade de responder ao paradoxo etiológico, bem como a de se criar Hipóteses Etiológicas Unificadas se esvazie, afinal, o que desponta é precisamente uma concepção anti-essencialista da etiologia da síndrome.
Dito isto, ao invés de potencialmente de contribuir para que se encerrem as especulações, no nosso entender este aspecto reabre uma via para que novos estudos emerjam sob o mote de se sondar o potencial etiológico de variáveis mentais/psicológicas diversas, sob as determinações de disposições pré-dispositivas.
Figura 2. Algoritmo sumarizando o funcionamento do modelo Epigênico Estendido (Dias, A.M. e Rodrigues, A.L.)
É de se ter em vista que desde que a literatura aceitou em seu âmago a máxima de que a esquizofrenia é uma desordem de cunho biológico, o estudo das variáveis não-orgânicas para a precipitação da síndrome gradualmente se dissipou, o que – à luz do nosso modelo – é anti-natural.
Por exemplo: utilizando o search engine biomédico de caráter semântico de Medi da Universidade de Tóquio, cuja estrutura é __1__2 ___3 ___ (e que, portanto, pode ser usado para busca semânticas no PubMed para X causa Y) para estresse causa/ induz/precipita esquizofrenia, revela que apenas 28 dos 15.976.265 artigos indexados no Medi usam esta sentença. Uma visada sobre os mesmo mostra que a maioria envolve o conceito de estresse materno, ou seja, apresentam o fator de risco epigênico como linha de estudo principal.
Nosso ponto de vista é claramente favorável a tal perspectiva, mas sugere que mais estudos devam ser dirigidos aos efeitos na transição para a vida adulta, eventualmente por força das cascatas moleculares precipitadas pelo estresse desde as vias de atuação neurohormonais, encabeçadas pelo efeito da exposição prolongada ao cortisol sobre o balanço serotonérgico e noradrenérgico e, desde então, sobre genes. É preciso que se lance um olhar sobre os casos que podiam ter sido e não foram – e manifestações subclínicas em geral – com vistas á delimitação do perfil de risco dos diversos fatores psicológicos e ambientais que podem contribuir para a precipitação da condição sindrômica, para que novos programas de saúde preventiva possam ser desenvolvidos, já em sentido à prevenção.
Resumo
Contexto: A esquizofrenia é uma síndrome complexa, relacionada a genes e fatores de risco nãogenéticos. Estudos epidemiológicos famosos reportaram sua presença em todas as culturas e regiões geográficas. Neste sentido, Hipóteses Etiológicas Unificadas enfrentam o desafio de simultaneamente apresentar dados experimentais e demonstrar que os mesmos dão conta do perfil universal da síndrome. Objetivos: revisar sistematicamente as mais proeminentes Hipóteses Etiológicas Unificadas, assim como a distribuição semântica dos achados genéticos (mediante técnicas de data mining) e propor um novo modelo, baseado nos efeitos dinâmicos de caráter epigênico sobre a ativação genética no neuro-desenvolvimento e puberdade. Resultados: de maneira geral, as Hipóteses Etiológicas Unificadas contradizem os principais achados genéticos (que sugerem que a esquizofrenia esteja associada ao perfil de neurotransmissores como D-1 e a cascata Glutamato-NMDA); também em geral, achados genéticos se encontram espalhados por todo o genoma (tal como revelamos em um mapa topológico dos 3519 estudos no assunto). A chave para este estado de coisas complexo pode ser representada pela associação entre a perspectiva de que cada polimorfismo associado à esquizofrenia represente um fator de risco estatístico (i.e. aumentando o risco de instabilidades do desenvolvimento), enquanto cascatas moleculares de caráter epigênico e fatores de risco ambientais permaneçam atuantes, através da ativação de genes em períodos críticos.
Referências
1. Andreone, N., Tansella, M., Cerini, R., Versace, A., Rambaldelli, G., Perlini, C. et al. Cortical white-matter microstructure in schizophrenia: Diffusion imaging study. The British Journal of Psychiatry, 191 (2), (2007), 113-119. [ Links ]
2. Ashtari, M., Cottone, J., Ardekani, B.A., Cervellione, K., Szeszko, P.R., Wu, J., et al. Disruption of white matter integrity in the inferior longitudinal fasciculus in adolescents with schizophrenia as revealed by fiber tractography. Archives of General Psychiatry, 64 (11), (2007), 1270-1280. [ Links ]
3. Bartley, A.J., Jones, D.W., Torrey, E.F., Zigun, J.R. & Weinberger, D.R. Sylvian fissure asymmetries in monozygotic twins: a test of laterality in schizophrenia. Biological Psychiatry, 34 (12), (1993), 853-863. [ Links ]
4. Bassett, A.S., Bury, A., Hodgkinson, K.A. & Honer, W.G. Reproductive fitness in familial schizophrenia. Schizophrenia Research, 21 (3), (1996), 151-160. [ Links ]
5. Bembenek, A. Analysis of the seasonality of birth effect in schizophrenic patients in Poland with the application of the seasonal decomposition method. Psychiatria Polska, 40 (2), (2006), 219-231. [ Links ]
6. Berlim, M.T., Mattevi, B.S., Belmonte-de-Abreu, P. & Crow, T.J. The etiology of schizophrenia and the origin of language: overview of a theory. Comprehensive Psychiatry, 44 (1), (2003), 7-14. [ Links ]
7. Bersani, G., Pucci, D., Gherardelli, S., Conforti, F., Bersani, I., Osborn, J. F., et al. Excess in the spring and deficit in the autumn in birth rates of male schizophrenic patients in Italy: potential role of perinatal risk factors. Journal of Maternal-Fetal Neonatal Medicine, 19 (7), (2006), 425-431. [ Links ]
8. Brown, J.S. Jr Identification of candidate genes for schizophrenia based on natural resistance to infectious diseases. Acta Neuropsychiatrica, 15 (3), (2003), 108-114. [ Links ]
9. Buchsbaum, M. S., Tang, C. Y., Peled, S., Gudbjartsson, H., Lu, D., Hazlett, E. A., et al. MRI white matter diffusion anisotropy and PET metabolic rate in schizophrenia. Neuroreport, 9 (3), (1998), 425-430. [ Links ]
10. Cheng, J.Y., Ko, J.S., Chen, R.Y. & Ng, E.M. Meta-regression analysis using latitude as moderator of paternal age related schizophrenia risk: high ambient temperature induced de novo mutations or is it related to the cold? Schizophrenia Research, 99 (1-3), (2008), 71-76. [ Links ]
11. Crow, T.J. Does schizophrenia have a cause? Trends in Cognitive Sciences, 10 (11), (2006), 478-479. [ Links ]
12. Crow, T.J. The Big Bang Theory of the origin of psychosis and the faculty of language. Schizophrenia Research, 102 (1-3), (2008), 31-52. [ Links ]
13. Crow, T.J., Done, D.J. & Sacker, A. Cerebral lateralization is delayed in children who later develop schizophrenia. Schizophrenia Research, 22 (3), (1996), 181-185. [ Links ]
14. Davis, K.L., Stewart, D.G., Friedman, J.I., Buchsbaum, M., Harvey, P.D., Hof, P. R., et al. White matter changes in schizophrenia: evidence for myelin-related dysfunction. Archives of General Psychiatry, 60 (5), (2003), 443-456. [ Links ]
15. Eaton, W.W., Byrne, M., Ewald, H., Mors, O., Chen, C.-Y., Agerbo, E., et al. Association of schizophrenia and autoimmune diseases: linkage of Danish National Registers. American Journal of Psychiatry, 163 (3), (2006), 521-528. [ Links ]
16. Ellman, L.M., Yolken, R.H., Buka, S.L., Torrey, E.F. & Cannon, T.D. Cognitive functioning prior to the onset of psychosis: the role of fetal exposure to serologically determined infl uenza infection. Biological Psychiatry, 65 (12), (2009), 1040-1047. [ Links ]
17. Erlenmeyer-Kimling, L. Mortality rates in the offspring of schizophrenic. Parents and a physiological advantage hypothesis. Nature, 220 (5169), (1968), 798-800. [ Links ]
18. Fan, J.B., Zhang, C.S., Gu, N.F., Li, X.W., Sun, W.W., Wang, H.Y., et al. Catechol-O-Methyltransferase gene Val/Met functional polymorphism and risk of schizophrenia: a large-scale association study plus meta-analysis. Biological Psychiatry, 57 (2), (2005), 139-144. [ Links ]
19. Fruntes, V. & Limosin, F. Schizophrenia and viral infection during neurodevelopment: a pathogenesis model? Medical Science Monitor, 14 (6), (2008), RA71-77. [ Links ]
20. Gallagher, B.J. 3rd, Jones, B.J., McFalls, J.A. Jr. & Pisa, A.M. Schizophrenic subtype, seasonality of birth and social class: a preliminary analysis. European Psychiatry, 22 (2), (2007), 123-128. [ Links ]
21. Hoffman, R.E. & Dobscha, S.K. Cortical pruning and the development of schizophrenia: a computer model. Schizophrenia Bulletin, 15 (3), (1989), 477-490. [ Links ]
22. Huxley, J., Mayr, E., Osmond, H. & Hoffer, A. Schizophrenia as genetic morphism. Nature, 204 (495), (1964), 220-221. [ Links ]
23. Jablensky, A. & Sartorius, N. Is schizophrenia universal? Acta Psychiatria Scandinavica Supplement, 344, (1988), 65-70. [ Links ]
24. Jablensky, A., Sartorius, N., Ernberg, G., Anker, M., Korten, A., Cooper, J.E., et al. Schizophrenia: manifestations, incidence and course in different cultures. A World Health Organization ten-country study. Psychological Medicine Monograph Supplement, 20, (1992), 1-97. [ Links ]
25. Jordaan, E., Niehaus, D.J., Koen, L., Seller, C., Mbanga, I. & Emsley, R.A. Season of birth, age and negative symptoms in a Xhosa schizophrenia sample from the Southern Hemisphere. Australian and New Zealand Journal of Psychiatry, 40 (8), (2006), 698-703. [ Links ]
26. Kallel, L., Brunelin, J., Zghal, A., Labbane, R., Dalery, J., dAmato, T., et al. Summer birth and deficit schizophrenia in Tunisia. Psychiatry Research, 152 (2-3), (2007), 273-275. [ Links ]
27. Konrad, A. & Winterer, G. Disturbed structural connectivity in schizophrenia. Primary factor in pathology or epiphenomenon? Schizophrenia Bulletin, 34 (1), (2008), 72-92. [ Links ]
28. Krystal, J.H., DSouza, D.C., Mathalon, D., Perry, E., Belger, A. & Hoffman, R. NMDA receptor antagonist effects, cortical glutamatergic function, and schizophrenia: toward a paradigm shift in medication development. Psychopharmacology, 169 (3-4), (2003), 215-233. [ Links ]
29. Lim, K.O. Connections in schizophrenia. American Journal of Psychiatry, 164 (7), (2007), 995-998. [ Links ]
30. Mayringer, H. & Wimmer, H. No deficits at the point of hemispheric indecision. Neuropsychologia, 40 (7), (2002), 701-704. [ Links ]
31. McGlashan, T.H. & Hoffman, R.E. Schizophrenia as adDisorder of developmentally reduced synaptic connectivity. Archives of General Psychiatry, 57 (7), (2000), 637-648. [ Links ]
32. Messias, E., Mourao, C., Maia, J., Campos, J.P., Ribeiro, K., Ribeiro, L., et al. Season of birth and schizophrenia in northeast Brazil: relationship to rainfall. Journal of Nervous and Mental Diseases, 194 (11), (2006), 870-873. [ Links ]
33. Oken, R. J. & Schulzer, M. At Issue: Schizophrenia and rheumatoid arthritis: the negative association revisited. Schizophrenia Bulletin, 25 (4), (1999), 625-638. [ Links ]
34. Pliskas, S. Neurociência para o clínico de saúde mental. Porto Alegre: ArteMed, (2004). [ Links ]
35. Roy, K., Murtie, J.C., El-Khodor, B.F., Edgar, N., Sardi, S.P., Hooks, B.M., et al. Loss of erbB signaling in oligodendrocytes alters myelin and dopaminergic function. A potential mechanism for neuropsychiatric disorders. Proceedings of the National Academy of Sciences, 104 (19), (2007), 8131-8136. [ Links ]
36. Rusch, N., Corrigan, P.W., Wassel, A., Michaels, P., Olschewski, M., Wilkniss, S. & Batia, K. A stress-coping model of mental illness stigma: I. Predictors of cognitive stress appraisal. Schizophrenia Research, 110 (1-3), (2009), 59-64. [ Links ]
37. Sanders, A.R., Duan, J., Levinson, D.F., Shi, J., He, D., Hou, C., et al. No significant association of 14 candidate genes with schizophrenia in a large European ancestry sample: implications for psychiatric genetics. American Journal of Psychiatry, 165 (4), (2008), 497-506. [ Links ]
38. Sawa, A. & Kamiya, A. Elucidating the pathogenesis of schizophrenia. British Medical Journal, 327 (7416), (2003), 632-633. [ Links ]
39. Schlosser, R.G., Nenadic, I., Wagner, G., Gullmar, D., von Consbruch, K., Kohler, S., et al. White matter abnormalities and brain activation in schizophrenia: a combined DTI and fMRI study. Schizophrenia Research, 89 (1-3), (2007), 1-11. [ Links ]
40. Shaner, A., Miller, G. & Mintz, J. Schizophrenia as one extreme of a sexually selected fitness indicator. Schizophrenia Research, 70 (1), (2004), 101-109. [ Links ]
41. Wilson, E.O. Da natureza humana. São Paulo: Queiroz Editores, (1981). [ Links ]
42. Yeo, R.A., Gangestad, S.W., Edgar, C. & Thoma, R. The evolutionary genetic underpinnings of schizophrenia: the developmental instability model. Schizophrenia Research, 39 (3), (1999), 197-206. [ Links ]
43. Yudofsky, S.C. Contracting schizophrenia: lessons from the infl uenza epidemic of 1918-1919. Journal of the American Medical Association, 301 (3), (2009), 324-326. [ Links ]
44. Zanetti, M.V., Schaufelberger, M.S., de Castro, C.C., Menezes, P.R., Scazufca, M., McGuire, P.K., et al. White-matter hyperintensities in first-episode psychosis. The British Journal of Psychiatry, 193 (1), (2008), 25-30. [ Links ]
45. Zhou, Y., Shu, N., Liu, Y., Song, M., Hao, Y., Liu, H., et al. Altered resting-state functional connectivity and anatomical connectivity of hippocampus in schizophrenia. Schizophrenia Research, 100 (1-3), (2008), 120-132. [ Links ]